Introduction
Vitamin K epoxide reductase (VKOR) is the enzyme responsible for regenerating vitamin K from vitamin K epoxide to support blood coagulation.
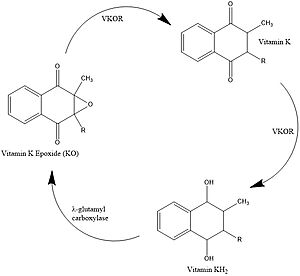
Figure 1. Vitamin K Cycle λ-glutamyl carboxylase uses Vitamin KH2 to carboxylate blood clotting cofactors, converting KH2 to KO in the process. Prothrombin is shown as an example of a blood clotting cofactor that later enters the coagulation cascade. VKOR converts KO back to KH2 via two steps with Vitamin K as an intermediate.
Vitamin K Cycle
Vitamin K is essential for blood clotting in the body[1]. The fully reduced form, KH2, is used by gamma-glutamyl carboxylase to carboxylate protein-bound glutamate residues in blood clotting cofactor precursors [2]. After carboxylation, the clotting cofactors (such as prothrombin) can bind to calcium and can proceed to the coagulation cascade [3]. During this process, KH2 becomes oxidized to Vitamin K epoxide, or KO [2]. VKOR turns the epoxide back to the fully reduced form so the reduced form can be used again. This transformation happens in two steps: 1) converting the epoxide to the partially oxidized Vitamin K quinone and 2) converting the quinone to the fully reduced hydroquinone (KH2) (Figure 1) [1].
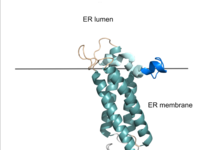
Figure 2. VKOR in Membrane The teal helices are the transmembrane helices. The tan sections are the beta loop and cap loop. The light blue section is the cap helix, and the dark blue section is the anchor. The cap loop, cap helix, and anchor make up the cap domain.
Structural Overview
VKOR consists of four embedded in the endoplasmic reticulum membrane. The barrel domain was used experimentally to stabilize VKOR for structure determination (Figure 2)[4]. For this page, the barrel domain has been removed and structures renumbered to correspond with the article by Liu. [4]. Helices one and two are connected by the region which contains two of the active cysteines, C43 and C51; these cysteines, along with C132 and C135, are essential for reduction and structural changes discussed in the next section[4]. VKOR also has a covering the active site, made up of an , , and . The anchor serves to attach the cap domain to the ER membrane for stabilization[4]. The loop helps stabilize one of the catalytic amino acids, Asn80[4]. The helix is involved in stabilization of certain disulfide bonds and structural changes as part of the catalytic cycle discussed below[4].
Active Site
VKOR uses two substrate binding amino acids, to stabilize vitamin K in the binding pocket. Tyr139 and Asn80 hydrogen bond to carbonyl groups on vitamin K and stabilizes it within the binding pocket [4]. Vitamin K is also bound via hydrophobic interactions within the binding pocket of VKOR. Hydrophobic residues of VKOR such as , form a hydrophobic tunnel within the binding pocket [4].
Catalytic Cycle
Catalytic Cysteines
The catalytic cycle of VKOR includes transitions from open to closed conformations by means of disulfide bridge-induced conformational changes (Figure 3). Open conformations (I & II) of VKOR exist when there is no ligand within the binding pocket. Closed conformations (III & IV) exist when some substrate exists within the binding pocket of VKOR. The substituent cysteines (I) act as reducing agents for the substrate, which can be either Vitamin K epoxide (KO) or partially reduced Vitamin K.
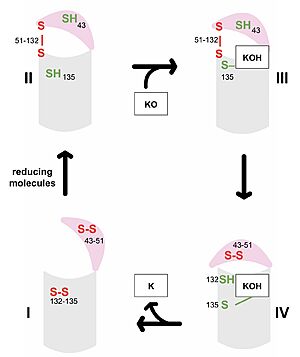
Figure 3. Catalytic Cycle of VKOR VKOR's luminal domain is represented by a the pink semicircle and the transmembrane domain is represented by the gray cylinder. Step I and II represent open conformations of VKOR and steps III and IV represent closed conformations.
The first step of the catalytic cycle (Figure 3) is the wild type open conformation. This step is characterized by an open cap domain with disulfide bonds between cysteines 43 and 51 and between cysteines 132 and 135 [4]. The second step of the catalytic cycle is a partially oxidized open conformation. This step is characterized by a disulfide bond between the luminal and transmembrane domain (Fig 3, step II). The transmembrane domain contains a free Cys135 and the luminal domain contains a free Cys43 [4]. Step II is labeled as open because no ligand exists within its binding pocket despite the disulfide bridge that connects the luminal and transmembrane domains. The next step of the cycle is a closed structure with an intact disulfide bond between Cys51 and Cys132. Cys135 is not involved in a disulfide bridge and instead reacts with substrate by forming a stable adduct with KOH or K. This binding induces the closed conformation and uses Cys43 in the luminal membrane for electron transfer [4]. The final step of the catalytic cycle is the last closed conformation. The Cys51-Cys132 bond is broken as Cys43 bonds with Cys51, recreating the disulfide bridge pattern of the open state. Cys132 is then free to bond with Cys135, releasing the product that was bound to the Cys135. Overall the catalytic cycle of VKOR is dependent on open and closed conformational changes of the protein and ultimately is used to generate vitamin K from vitamin K epoxide [4].
Medical Relevance
Warfarin
Warfarin is the most widely prescribed oral anticoagulant and targets blood clotting via inhibition of VKOR. The FDA approved uses for cardiac conditions (myocardial infarction, atrial fibrillation) as well as for deep vein thrombosis and pulmonary embolism. Due to the inhibition of the normal blood clotting cycle, patients taking warfarin are at risk for hemorrhage which can occur anywhere in the body. [5]
Warfarin is a of Vitamin K that occupies the VKOR binding site, acting as a competitive inhibitor. Warfarin mimics vitamin K by binding to the same (Asn80 and Tyr139) in the active site. Warfarin also shares the same within the binding site that KOH experiences (Phe83, Phe87, and Tyr88). Warfarin binding also depends on the VKOR catalytic cysteines. Warfarin is able to bind to the fully oxidized open form of VKOR as shown in of the catalytic cycle. Once Warfarin binds, VKOR is considered to be in a closed conformation since the substrate cannot enter, despite the lack of disulfide bridge changes. Warfarin can also bind to the partially oxidized form of VKOR as shown in of the catalytic cycle.
There are around 30 known missense mutations that lead to warfarin resistance in patients, but these mutations do not affect Vitamin K binding for reasons which are not yet fully understood. Such patients require higher doses of warfarin to reach therapeutic level or require a different anticoagulant drug. [6]
Superwarfarins
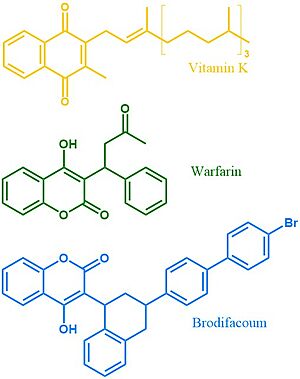
Figure 4. Vitamin K, Warfarin, and Brodifacoum Above is a comparison of the 2D structures of VKOR's natural substrate, the blood thinner warfarin, and the superwarfarin brodifacoum.
More potent warfarin derivatives, called superwarfarins, are used as rodenticides. Superwarfarins have bulkier side chains that allow them to stay bound to VKOR for long periods of time, causing prolonged and uncontrolled bleeding[7]. The enhanced binding is due to interactions between the bulky side chains and the hydrophobic residues of VKOR's binding pocket. The duration of one superwarfarin, brodifacoum, has been reported as 15-30 days [7] vs. the clinical warfarin duration of 2-5 days[5].




