This old version of Proteopedia is provided for student assignments while the new version is undergoing repairs. Content and edits done in this old version of Proteopedia after March 1, 2026 will eventually be lost when it is retired in about June of 2026.
Apply for new accounts at the new Proteopedia. Your logins will work in both the old and new versions.
Sandbox Reserved 466
From Proteopedia
Contents |
TETANUS TOXIN
| This Sandbox is Reserved from 13/03/2012, through 01/06/2012 for use in the course "Proteins and Molecular Mechanisms" taught by Robert B. Rose at the North Carolina State University, Raleigh, NC USA. This reservation includes Sandbox Reserved 451 through Sandbox Reserved 500. | ||||||
To get started:
More help: Help:Editing For more help, look at this link: http://www.proteopedia.org/wiki/index.php/Help:Getting_Started_in_Proteopedia IntroductionThe tetanus toxin is produced by the bacteria Clostridium tetani. Clostridium bacteria produces 8 distinct neurotoxins that are extremely potent to humans. This spore-forming bacillus bacteria is widely found in nature, particularly in soil. It enters the body through cuts or abrasion of the skin. The Clostridium bacteria produces two types of neurotoxins. Both Clostridium botulinum and Clostridium tetani form the clostridial neurotoxin family [Rao et al., 2005] and have high homology between them. This family is classified as part of the endopeptidase M27 family of proteins, which are metalloproteases. Metalloproteases bind with a divalent cation, usually zinc, which activates water molecules within the active site to hydrolyze peptide bonds [PDB]. This neurotoxin is catalytically classified as a hydrolase. The active site forms a nucleophilic water, which cleaves a peptide bond of the substrate.
Structure
The 150 kDa precursor polypeptide of the tetanus toxin is cleaved during post-translational modification into a heavy and light chains. These two chains remain linked by a disulfide bridge. The 100 kDa heavy chain is the C-terminal end of the protein and the 50 kDa light chain is the N-terminal end of the protein. If the two chains are separated, the toxin becomes non-toxic [PDB].
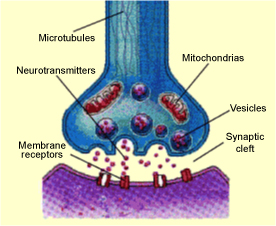
Mechanism of ActionThe tetanus toxin acts in the synaptic cleft of neuronal cells and prevents the release of neurotransmitters from the presynaptic neuron terminal [Link et al., 1992]. Neurotransmitters are released from the presynaptic terminal of a neuron into the synaptic cleft and received by endocytosis by the postsynaptic terminal of the next neuron. Nerve terminals are filled with vesicles, which are specialized storage components that contain neurotransmitters, and are released from the terminal into the synaptic cleft by exocytosis. The surface of the postsynaptic terminal has many specialized receptors, which bind with specific neurotransmitters. The neurotransmitters are then endocytized into the postsynaptic neuron for transmission of the synapse to facilitate a response to the nerve stimuli [Dasgupta, 1993].
Medical Implications or Possible ApplicationsThe tetanus toxin effects the central nervous system by inhibiting the release of neurotransmitters, glycine and gamma-aminobutyric acid, into the synaptic cleft of the spinal cord affecting the transmission of nerve impulses throughout the body. This toxin causes tetanus, which is characterized by rigidity, spasms, and paralysis of the voluntary muscles of the body [Rao et al., 2005]. Tetanus is often referred to as "lockjaw" because a large majority of the patients experience rigidity of the jaw muscles. The toxin enters the body by way of a cut and enters the bloodstream, where it spreads rapidly throughout the body, or by a nerve, which transports the toxin directly to the central nervous system. Tetanus toxin attacks motor nerve cells and hyper-activates them. The overactive nerve impulses cause muscles to go into convulsive spasms. The toxin is most commonly known to affect the muscles of the jaw causing rigidity of the muscles of the jaw and face. This toxin also causes severe spasms in the throat and chest making swallowing and breathing extremely difficult. These are the most common causes of death if tetanus is untreated. Tetanus also causes adverse effects on various muscles throughout the body, notably on the heart, blood pressure, and vital brain centers that cause death later in the disease [Montecucco, 1995].
ReferencesBizzini, B. (1979). Tetanus toxin. Microbiology and Molecular Biology Reviews, 43(2), 224-240. Retrieved from http://mmbr.asm.org/content/43/2/224. Caccin, P., Rossetto, O., Rigoni, M., & Johnson, E. (2003). Vamp/synaptobrevin cleavage by tetanus and botulinum neurotoxins is strongly enhanced by acidic liposomes. FEBS Letters, 542(1-3), 132-136. Retrieved from http://www.sciencedirect.com/science/article/pii/S001457930300365X. Dasgupta, B. R. (1993). Botulinum and tetanus neurotoxins: Neurotransmission and biomedical aspects. New York: Plenum Press. Link, E., Edelmann, L., Chou, J., & Binz, T. (1992). Tetanus toxin action: Inhibition of neurotransmitter release linked to synaptobrevin proteolysis. Biochemical and Biophysical Research Communications, 189(2), 1017-1023. Retrieved from http://www.sciencedirect.com/science/article/pii/0006291X9292305H. Montecucco, C. (1995). Clostridial neurotoxins: The molecular pathogenesis of tetanus and botulism. Germany: Springer. Rao, K. N., Kumaran, D., Binz, T., & Swaminathan, S. (2005). Structural analysis of the catalytic domain of tetanus neurotoxin. Microbiology and Molecular Biology Reviews, 45, 929-939. Retrieved from www.elsevier.com/locate/toxicon. Rossetto, O., Caccin, P., Rigoni, M., & Tonello, F. (2001). Active-site mutagenesis of tetanus neurotoxin implicates tyr-375 and glu-271 in metalloproteolytic activity. Toxicon, 39(8), 1151-1159. Retrieved from http://www.sciencedirect.com/science/article/pii/S004101010000252X. Rutgers, & UCSD. (2012, May 1). 1yvg. Retrieved from Protein Data Base website: http://www.rcsb.org/pdb/explore/explore.do?structureId=1yvg#. |

{kind=link}
{kind=link}
{kind=link}