Myosin is a large asymmetric molecule with a MW of about 500,000 kDa. It consist of two globular head domains termed myosin subfragment 1 (S1), one neck subfragment 2 (S2) and a light meromyosin tail (LMM) [3]. Myosin S1 unit comprises of a motor domain (MD) and a lever arm (Fig.3) By 2000 scallop myosin S1 has been determined in three different conformations of the contractile cycle, corresponding to the following structures [4]:
• S1 nucleotide-free state corresponding to the near-rigor conformation of myosin 1DFK
• S1 Mg-ADP • BeFx state corresponding to the pre-power stroke conformation 1KK8
• S1 Mg-ADP state corresponding to the myosin detached state 1B7T
Comparing the available crystal structures of different myosin S1 units enable us to understand the conformational changes within the motor domain during the contractile cycle. Here mainly the structure and function of the MD in the S1 Mg-ADP detached state will be discussed.
The subdomains of the motor domain
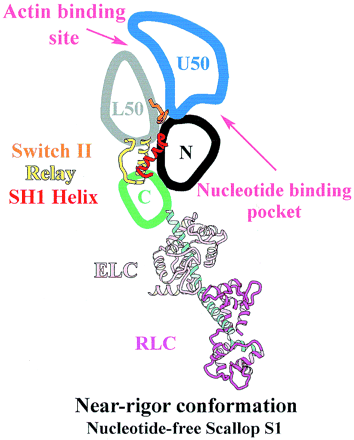
Figure 3. The subdomains on myosin S1 unit
[4]The MD of the scallop S1 unit is most frequently described as consisting of four subdomains: the converter, the N-terminal subdomain, and the upper and lower 50-kDa subdomains [5]. They are linked together by three single-stranded joints termed the switch II (residues IIe-461 to Asn-470), the relay (residues Asn-489 to Asp-519), and SH1 helix (residues Cys-693 to Phe-707) (Fig. 3) [4].
• Of the MD subdomains the converter changes its position the most during the contractile cycle. Connection of the converter and the lever arm allows relatively small changes in the converter to be greatly amplified in the lever arm (Fig. 4)
• Upper 50-kDa subdomain and N-terminal subdomain form the nucleotide binding pocket
• Lower and upper 50-kDa subdomains form the interface where actin can bind [4]
Conformational changes in the flexible joints coordinate rearrangements of the four MD subdomains enabling the transition between different myosin S1 conformations in the actomyosin contractile cycle, during which S1 traduces ATP hydrolysis to mechanical work. The different conformational states of myosin are termed strong or weak actin-binding states [6].
Nucleotide binding pocket: ADP + Mg2+
The nucleotide binding pocket is located at the interface of the 50 kDa upper subdomain and the N-terminal subdomain [6], which is opposite to a deep cleft that bisects the actin-binding domain (Fig. 3). The nucleotide binding pocket and actin binding domain contained a complex arrangement of secondary structure elements mainly around the parallel 7-stranded . Loops extending from the β-strands interact with the adenine nucleotide.
forms hydrogen bonds with the amino acid side chains in the binding pocket. coordinates with residues Thr183, Ser 241 of the N-terminal subdomain as well as O1B and O3B from ADP and three water molecules. The hydrogen bonds between ADP and the amino acid residues together with the interactions of Mg2+ keep ADP in the nucleotide-binding pocket.
In the contractile cycle ATP binding causes a conformational change, which detaches the myosin S1 unit from actin. Then the active site closes, and ATP is hydrolysed to Pi and ADP, leading to the subsequent reattachment of the S1 with the actin. The conformational changes of the acting-binding pocket and the opening and closing of the nucleotide-binding pocket cause the strong and weak acting binding states of myosin, allowing muscle contraction [4].
Role of the subdomains and joints in the mechanism of the contractile cycle
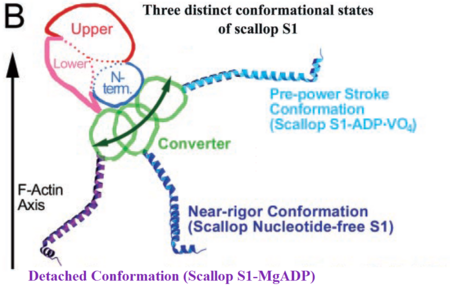
Figure 4. Relatively small movement of the converter is amplified by the lever arm (adaptation of
[7][4]).
The MD has different conformational states in each step of the contractile cycle. The conformation of the MD in each state depends on which nucleotide is bound to the active site (if any). In each structural state the conformation of the MD changes relatively little, but these changes are enough to cause a substantial difference in the position of the lever arm (Fig. 4) [4].
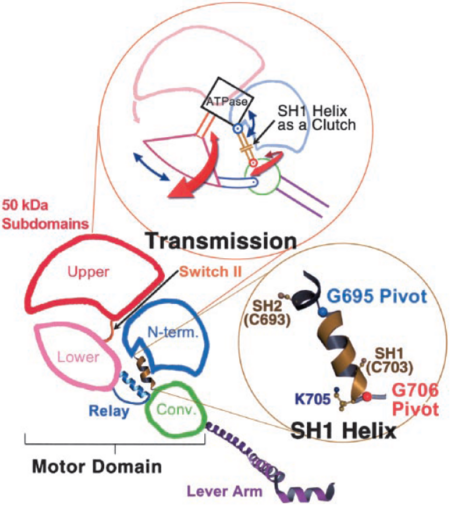
Figure 5. Arrows in the upper figure show the direction of rotation of the 50-kDa lower and upper subdomains and the converter around the N-terminal subdomain. Lower picture shows the pivots around which SH1 unwinds
[7].
The 50-kDa upper and lower subdomains as well as the converter control the motor function of the myosin head by rotating around the N-terminal subdomain (Fig 5). The rotations depend on the conformational changes of the 3 joints; switch II, SH1 helix region, and relay [6]. The joints work together in the transition between the different conformational states of MD to control the overall organization of the myosin head. They also allow communication between the nucleotide-bonding pocket, acting-binding interface and the lever arm [4].
Switch II, a catalytic loop of the nucleotide-binding pocket, moves in and out of the nucleotide-binding pocket during enzymatic activity. It is responsible for the unwinding of SH1 helix, along with the conformational changes caused by nucleotide binding. As SH1 unwinds, by rotating around two pivots (G695, G706, Fig. 5), it uncouples the converter/lever module from the MD [7]. Movement of the converter is controlled by the relay joint. The converter/relay module attains different conformations changing the position of the lever arm (Fig. 4) and thus giving rise to the different states in the actomyosin cycle [4].
Taking part in the organization of the different conformations of the contractile cycle is also the so called switch I, which is a second catalytic loop of the nucleotide-binding pocket. [4] Switch II forms a specific salt bridge and hydrogen bond interactions with switch I that stabilize the pre-power stroke state [7].
In the detached conformation of the MD, switch II interacts with the nucleotide-binding pocket and forms the stabilizing hydrogen bond interactions and a salt bridge with switch I [7][6]. Rotation of the 50-kDa upper subdomain away from the N-terminal subdomain pulls switches I and II apart breaking the protein- nucleotide interactions between switch I and ADP, as well as changing the conformation of switch II. These changes result in closing of the actin-binding site and opening of the nucleotide-binding pocket, leading to MgADP release. At the same time SH1 helix is unwound and the lever arm is able to change its position enabling sliding of myosin through the actin filament [7].
Conclusions
The conformational changes of the myosin head during the contractile cycle allow the myosin filaments to slide across the actin filaments in the muscle sarcomere, producing movement. Scallop myosin is an exceptional system in that it is the first myosin isoform that has crystallized in all the three states of the contraction cycle: the near-rigor state, pre-power stroke state and the detached state. However, not all conformational changes and interactions of the S1 with the nucleotides are known, even though kinetic studies and electron microscopy (EM) have helped to understand the mechanism of the contractile cycle along with the information from structural studies. Future perspectives for muscle and myosin research are to characterize a structure with S1 bound to actin, elucidate the structural details of how nucleotide binding is coupled to actin affinity, and to define the various conformational states of myosin in species other than scallop.





