The RNase H2 ribonuclease complex is a heterotrimeric endoribonuclease responsible for the major ribonuclease H activity in mammalian cells. In mouse, the complex is encoded by 3 genes located on chromosomes 8 (Rnaseh2a), 14 (Rnaseh2b) and 19 (Rnaseh2c)[1]. This enzyme specifically cleaves the 3’O-Phosphate bond of RNA in a DNA/RNA hybrids to produce 5’ phosphate and 3’hydroxyl ends.
Biological role
Ribonucleases H are the only known enzymes, able to degrade the RNA strand of a DNA/RNA hybrid in a sequence-nonspecific way.
There are two types of RNase H (RNases H1 and RNases H2) classified according to their sequence conservation and substrate preference. Currently, three types of RNA/DNA hybrids are known: simple RNA/DNA duplexes (Figure 1A), RNA•DNA/DNA hybrids (Figure 1B), and DNA•RNAfew•DNA/DNA hybrids (Figure 1C). RNases H2 is totally able to cleave a single ribonucleotide embedded in a double strand DNA (DNA• RNAfew •DNA/DNA type) when RNases H1 require at least 4 ribonucleotides. This ability and their high expression in proliferating cells suggest that RNases H2 are involved in DNA repair and replication[2].

Figure 1 : Three types of RNA/DNA hybrids
Indeed, ribonucleotides are wrongly incorporated into DNA during DNA replication at a frequency of about 2 ribonucleotides per kb. With such frequency, these errors are by far the most abundant threat of DNA damaging. Hence, a correction is essential to the preservation of DNA integrity: the most common correction mechanism involves RNases H2 and is called Ribonucleotide Excision Repair (RER). The incorporation of ribonucleotides in DNA produce DNA•RNAfew•DNA/DNA hybrids from which the few misincorporated ribonucleotides can be removed by an RNase H2[3].
This repair activity is guided by the interaction between C-terminus of RNase H2B protein and the DNA clamp PCNA. This interaction occurs through a hydrophobic conserved peptide motif called the PCNA interaction peptide PIP (PIP-box: Residues 294 to 301 MKSIDTFF of H2B protein) that interacts with a hydrophobic groove near the PCNA C-terminus. This interaction allows RNase H2 to scan DNA for misincorporated ribonucleotides which makes the Ribonucleotide Excision Repair more efficient[4].
Furthermore, in vitro studies have shown that RNases H2 is likely to be involved in the removal of RNA primer from Okazaki fragment produced during the synthesis of the lagging strand in DNA replication since Okazaki fragment are RNA•DNA/DNA hybrids (Figure 1B)[4].
RNases H2 activity is crucial in mammalian cells, for instance a mutation in human RNase H2 causes Aicardi-Goutières syndrome. This syndrome is an auto-inflammatory disorder that may be the consequence of an increased production of incorrect nucleic acid by-products during DNA replication[4].
Structure
A heteromeric complex
It has been shown that the Mammalian RNase complex is a heteromeric complex formed by 3 distinct proteins: , and . H2A protein is the catalytic subunit and H2B/H2C proteins are auxiliary subunits: they are structural domains that facilitate cohesion of the complex[5].
The first domain structure of the complex, H2A, contains 301 amino acids, almost as H2B protein which computes 308 amino acids. H2C protein is the smallest subunit: it only has 166 amino acids.
Each of these proteins adopts various with β-strands and α-helices:
- H2A protein has 12 α-helices, 11 β-strands and 3 turns[6],
- H2B molecule computes 8 α-helices, 7 β-strands and 3 turns[7],
- H2C subunit consists of 5 α-helices, 8 β-strands and 2 turns[8].
Several interactions between the subunits
H2C protein is found in the middle of the elongated complex structure, flanked by H2A and H2B proteins on the ends.
The complex is stabilized by the intimately interwoven architecture of H2B and H2C: The N-terminal region of H2B protein (amino acids 1-92) weaves together with H2C domain to form 3 β-barrels, called [9]. This triple barrel is formed from a total of and produces a pseudo-2-fold axis of symmetry along the central barrel. Also, it permits to leave the mostly α-helical C-terminal region of H2B available for potential interactions with other protein (for example the PCNA protein). Finally, it has been found that the motif provides a platform for securely binding the H2A protein: the side and end of the first barrel in the subcomplex H2B/H2C form a with amino acids 197-258 in the C-terminal region of H2A protein. This interface is mainly composed of hydrophobic residues[5]
Interactions with nucleic acids
It has been proved that the position of RNA/DNA complex in the active site cleft is determined by several favorable electrostatic interactions between the nucleic acid and positively charged amino acids of the protein[2].
The β6-α6 loop of the H2A protein could play a role in substrate recognition: the minor groove of the double helix molecule straddles this area of the protein, which results in a non-sequence specific cleavage by the enzyme. Moreover, the β6-α6 loop contains a , which might act as a sensor for the hybrid by forming an interaction with the 2’-hydroxyl group of the ribose in the 3’ nucleotide of the RNA primer in the RNA-DNA hybrid .Another Lysine might be involved in the recognition of nucleic acids, indeed might be able to recognize the 2'-hydroxyl groupe of the ribose of the amino acid at 5'end of the RNA (Figure 2)[5]. Therefore, since DNA does not contain a 2’-hydroxyl group in it nucleotide sequence, the RNase H2 can only recognize RNA in the hybrid: only ribonucleotides of the RNA strand are positioned in the active site. The RNA-DNA hybrid is placed such that the target phosphodiester bond between the RNA and DNA parts of the hybrid is in the proper orientation for nucleophile attack by a two-metal ion mechanism.
It is important to notice that the Mammalian RNase H2 contains only one cleft with the active site for substrate binding: RNase H2 may recognize single ribonucleotide within a DNA duplex that have a B-form helical structure, as well as longer RNA in RNA-DNA hybrid which adopts intermediate A/B form structure[9]. Thus, the RNase H2 enzyme needs to bind both conformations to able to fully complete all its roles.
Activity
The RNase H2 recognize 2’OH group of ribonucleotides in RNA at RNA/DNA junction and cannot cleave unhybridized RNA. The phosphodiester hydrolysis catalysed by the RNase H2 is likely following a two-metal ion-dependent mechanism quite common for phosphoryl hydrolases like RNase H enzymes.
The active site of the catalytic H2A protein is located in a cleft near one end of the complex and formed by strands β2 and β5, and helices α4 and α5. The active site contains four catalytic amino acids strictly conserved in RNases H2: [5]. These amino acids are essential for the coordination of the 2 divalent metal ions implicated in the stabilisation of reaction intermediates and transition states. In vitro, these ions can be Mg++, Mn++ or Zn++ but the native enzyme is likely to contain Mg++[10].
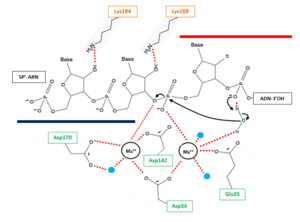
Figure 2 : Proposed mechanism for Okazaki fragment processing (not a concerted mechanism)
The hydrolysis can be decomposed in 3 steps:
- 1 : Deprotonation of a water molecule coordinated to the metal MB++ to form a nucleophile OH- ion. This hydroxide ion will then be properly oriented for an in-line nucleophilic attack of the target phosphate. The deprotonation mechanism has not been elucidated yet but two hypothesis can explain this step. According the first one, the metal MB++ might be responsible for the generation of water nucleophile. The other one involve a participation of the pro-R oxygen of the phosphate immediately to the 3’ side of the scissile bond, which is likely to serve as a general base for deprotonation (with transfer of the H+ to the solvent).This pro-R oxygen is also thought to play a role in the proper orientation of the hydroxide ion[11].
- 2 : In line attack by hydroxide ion of the target phosphate. During this step, a pentacovalent phosphate (transition state) is formed and stabilized by metal MA++. The metal MA++ interacts with both the nonbridging and 3' bridging oxygen and the strain induced by this double briding is thought to accelerate the hydrolysis rate. This step release a 3’ oxyanion stabilized by metal MA++ acting as a Lewis acid[10].
- 3 : The cleaved phosphate cannot simultaneously coordinate the two metal ions anymore, and likely one of the metal ions leave the active site which triggers a release of cleave product[9].


