This old version of Proteopedia is provided for student assignments while the new version is undergoing repairs. Content and edits done in this old version of Proteopedia after March 1, 2026 will eventually be lost when it is retired in about June of 2026.
Apply for new accounts at the new Proteopedia. Your logins will work in both the old and new versions.
User:Karen Lee/Sandbox 1
From Proteopedia
(→Tus Polar Fork Arrest) |
|||
| Line 1: | Line 1: | ||
| - | = | + | ='''Replication Termination'''= |
| - | Replication is an essential process in all cells. The process copies the chromosomal DNA of the organism to provide the extra copy needed in cell division and is therefore critical in the biological inheritance of genes. In cells with circular chromosomes, replication starts from a single origin proceed with two replication forks moving in opposite directions. | + | Replication is an essential process in all cells. The process copies the chromosomal DNA of the organism to provide the extra copy needed in cell division and is therefore critical in the biological inheritance of genes. In cells with circular chromosomes, replication starts from a single origin proceed with two replication forks moving in opposite directions. This process must be terminated, otherwise multiple copies of the chromosome would be made. |
The process of terminating replication is performed by replication termination proteins. These proteins bind to specific sequences in the DNA, called ''Ter'' sites. This binding provides a physical blockage in the DNA that stops the replication machinery. In ''B. subtilis'', the termination protein is called Replication Terminator Protein (RTP), and in ''E. coli'' it is Termination Utilisation Substance (Tus). | The process of terminating replication is performed by replication termination proteins. These proteins bind to specific sequences in the DNA, called ''Ter'' sites. This binding provides a physical blockage in the DNA that stops the replication machinery. In ''B. subtilis'', the termination protein is called Replication Terminator Protein (RTP), and in ''E. coli'' it is Termination Utilisation Substance (Tus). | ||
| - | + | The replication terminator proteins function by binding to specific DNA sequences called''Ter'' sites. Each circular chromosome has two sets of ''Ter'' sites that appear roughly opposite to the origin of replication. One set blocks the clockwise replication fork while the other traps the anti-clockwise replication fork. Interestingly, both these proteins bind to DNA in a polar manner that blocks the replication fork in one direction, but allow it to continue in when approached from the other direction. | |
| - | ==RTP== | + | =='''RTP'''== |
<Structure load='1f4k' size='400' frame='true' align='right' caption='An RTP dimer bound to the B-site Ter sequence' scene='Insert optional scene name here' /> | <Structure load='1f4k' size='400' frame='true' align='right' caption='An RTP dimer bound to the B-site Ter sequence' scene='Insert optional scene name here' /> | ||
| - | ===Structure of RTP=== | + | ==='''Structure of RTP'''=== |
A single RTP monomer consists of four <scene name='User:Karen_Lee/Sandbox_1/Rtp_alpha_helices/3'>α-helices</scene>, three <scene name='User:Karen_Lee/Sandbox_1/Rtp_beta_sheets/2'>β-strands</scene> and an unstructured <scene name='User:Karen_Lee/Sandbox_1/Rtp_n_terminal/1'>N-terminal</scene> domain. The <scene name='User:Karen_Lee/Sandbox_1/Rtp_alpha_helix_3_dna/3'>α3 helix</scene> binds to DNA by <scene name='User:Karen_Lee/Sandbox_1/Rtp_alpha_helix_3_dna/1'>inserting</scene> into the major groove of Ter sites, while the <scene name='User:Karen_Lee/Sandbox_1/Rtp_beta_strand_dna/1'>β2-strand</scene> interacts with the minor groove. The N-terminal arm also binds to the Ter site<ref>Pai, S. K., Bussiere, D. E., Wang, F., Hutchinson, C. A., White, S. W. & Bastia, D. (1996) The structure and function of the replication terminator protein of Bacillus subtilis: identification of the ‘winged helix’ DNA-binding domain. ''EMBO J.'' '''15(12)''', 3164-3173.</ref>. | A single RTP monomer consists of four <scene name='User:Karen_Lee/Sandbox_1/Rtp_alpha_helices/3'>α-helices</scene>, three <scene name='User:Karen_Lee/Sandbox_1/Rtp_beta_sheets/2'>β-strands</scene> and an unstructured <scene name='User:Karen_Lee/Sandbox_1/Rtp_n_terminal/1'>N-terminal</scene> domain. The <scene name='User:Karen_Lee/Sandbox_1/Rtp_alpha_helix_3_dna/3'>α3 helix</scene> binds to DNA by <scene name='User:Karen_Lee/Sandbox_1/Rtp_alpha_helix_3_dna/1'>inserting</scene> into the major groove of Ter sites, while the <scene name='User:Karen_Lee/Sandbox_1/Rtp_beta_strand_dna/1'>β2-strand</scene> interacts with the minor groove. The N-terminal arm also binds to the Ter site<ref>Pai, S. K., Bussiere, D. E., Wang, F., Hutchinson, C. A., White, S. W. & Bastia, D. (1996) The structure and function of the replication terminator protein of Bacillus subtilis: identification of the ‘winged helix’ DNA-binding domain. ''EMBO J.'' '''15(12)''', 3164-3173.</ref>. | ||
| Line 17: | Line 17: | ||
(<scene name='User:Karen_Lee/Sandbox_1/Rtp_revert/1'>Revert to original view</scene>) | (<scene name='User:Karen_Lee/Sandbox_1/Rtp_revert/1'>Revert to original view</scene>) | ||
| - | ===RTP Polarity=== | + | ==='''RTP Polarity'''=== |
The functional RTP-DNA complex requires two dimers of RTP. Dimerisation occurs through the extended <scene name='User:Karen_Lee/Sandbox_1/Alpha4_helix_interaction/1'>α4-helix</scene> of each monomer forming an anti-parallel coiled coil. The RTP dimer binds to the B site first which then facilitates cooperative binding of another RTP dimer to the A site to form a complete RTP-Ter complex<ref name="Vivian" />. | The functional RTP-DNA complex requires two dimers of RTP. Dimerisation occurs through the extended <scene name='User:Karen_Lee/Sandbox_1/Alpha4_helix_interaction/1'>α4-helix</scene> of each monomer forming an anti-parallel coiled coil. The RTP dimer binds to the B site first which then facilitates cooperative binding of another RTP dimer to the A site to form a complete RTP-Ter complex<ref name="Vivian" />. | ||
| Line 28: | Line 28: | ||
As RTP binds asymmetrically across the nRB, due to its differing binding affinities for the B and A sites, a polar complex is formed to block the replication fork. When the fork approaches the B site where there is tight RTP-DNA binding, the fork is unable to progress. But approaching from the A site, where there is a lower binding affinity, the fork is able to pass the complex. | As RTP binds asymmetrically across the nRB, due to its differing binding affinities for the B and A sites, a polar complex is formed to block the replication fork. When the fork approaches the B site where there is tight RTP-DNA binding, the fork is unable to progress. But approaching from the A site, where there is a lower binding affinity, the fork is able to pass the complex. | ||
| - | ===RTP as a checkpoint regulator=== | + | ==='''RTP as a checkpoint regulator'''=== |
An interesting feature is that RTP can cause replication fork pausing in ''B. subtilis'' at regions other than ''Ter'' sites, acting as a replication checkpoint regulator. There appear to be stringent terminator (''STer'') regions where RTP can bind and arrest replication when cells are under stress i.e. the stringent response. These regions are located around 200 kb either side of the origin of replication<ref>Levine, A., Vannier, F., Dehbi, M., Henckes, G. & Seror, S. J. (1991) The stringent response blocks DNA replication outside the ''ori'' region in ''Bacillus subtilis'' and at the origin in ''Escherichia coli''. ''J. Mol. Biol.'' '''219''', 605-613.</ref>. Studies have shown that there is 76% homology between this site and the regular ''Ter'' sites<ref>Autret, S., Levine, A., Vannier, F., Fujita, Y. & Seror, S. J. (1999) The replication checkpoint control in ''Bacillus subtilis'': identification of a novel RTP-binding sequence essential for the replication fork arrest after induction of the stringent response. ''Mol. Microbiol.'', '''31''', 1665-1679.</ref>. Additionally, mutational studies have reported that interaction between two dimers may not be required for arrest at ''STer'' sites. This is not the case with regular ''Ter'' sites. | An interesting feature is that RTP can cause replication fork pausing in ''B. subtilis'' at regions other than ''Ter'' sites, acting as a replication checkpoint regulator. There appear to be stringent terminator (''STer'') regions where RTP can bind and arrest replication when cells are under stress i.e. the stringent response. These regions are located around 200 kb either side of the origin of replication<ref>Levine, A., Vannier, F., Dehbi, M., Henckes, G. & Seror, S. J. (1991) The stringent response blocks DNA replication outside the ''ori'' region in ''Bacillus subtilis'' and at the origin in ''Escherichia coli''. ''J. Mol. Biol.'' '''219''', 605-613.</ref>. Studies have shown that there is 76% homology between this site and the regular ''Ter'' sites<ref>Autret, S., Levine, A., Vannier, F., Fujita, Y. & Seror, S. J. (1999) The replication checkpoint control in ''Bacillus subtilis'': identification of a novel RTP-binding sequence essential for the replication fork arrest after induction of the stringent response. ''Mol. Microbiol.'', '''31''', 1665-1679.</ref>. Additionally, mutational studies have reported that interaction between two dimers may not be required for arrest at ''STer'' sites. This is not the case with regular ''Ter'' sites. | ||
| - | ==Tus== | + | =='''Tus'''== |
<Structure load='1ecr' size='400' frame='true' align='left' caption='A TUS monomer complexed with DNA' /> | <Structure load='1ecr' size='400' frame='true' align='left' caption='A TUS monomer complexed with DNA' /> | ||
| Line 39: | Line 39: | ||
(<scene name='User:Karen_Lee/Sandbox_1/Tus_reset/1'>Revert to original view</scene>) | (<scene name='User:Karen_Lee/Sandbox_1/Tus_reset/1'>Revert to original view</scene>) | ||
| - | ==Tus Polar Fork Arrest Mechanism== | + | ==='''Tus Polar Fork Arrest Mechanism'''=== |
Like the RTP Ter sites, the Tus ''Ter'' sites do not display any symmetry or inverted repeats, thus a polar complex is formed between the Tus protein and ''Ter'' site. Despite both being replication termination proteins, RTP and Tus have very different structures and mechanisms of polar fork arrest. | Like the RTP Ter sites, the Tus ''Ter'' sites do not display any symmetry or inverted repeats, thus a polar complex is formed between the Tus protein and ''Ter'' site. Despite both being replication termination proteins, RTP and Tus have very different structures and mechanisms of polar fork arrest. | ||
| Line 53: | Line 53: | ||
The unlocking of the complex after it has performed it function of terminating replication from the non-permissive face, is ultimately required for the replication fork in the other direction to proceed. There is no specific unlocking mechanism, this is simply achieved as previously described, with the sliding movement of the DnaB helicase knocking Tus off its ''Ter'' sequence. | The unlocking of the complex after it has performed it function of terminating replication from the non-permissive face, is ultimately required for the replication fork in the other direction to proceed. There is no specific unlocking mechanism, this is simply achieved as previously described, with the sliding movement of the DnaB helicase knocking Tus off its ''Ter'' sequence. | ||
| - | ==References== | + | =='''References'''== |
<references /> | <references /> | ||
Revision as of 12:06, 16 May 2011
Contents |
Replication Termination
Replication is an essential process in all cells. The process copies the chromosomal DNA of the organism to provide the extra copy needed in cell division and is therefore critical in the biological inheritance of genes. In cells with circular chromosomes, replication starts from a single origin proceed with two replication forks moving in opposite directions. This process must be terminated, otherwise multiple copies of the chromosome would be made.
The process of terminating replication is performed by replication termination proteins. These proteins bind to specific sequences in the DNA, called Ter sites. This binding provides a physical blockage in the DNA that stops the replication machinery. In B. subtilis, the termination protein is called Replication Terminator Protein (RTP), and in E. coli it is Termination Utilisation Substance (Tus).
The replication terminator proteins function by binding to specific DNA sequences calledTer sites. Each circular chromosome has two sets of Ter sites that appear roughly opposite to the origin of replication. One set blocks the clockwise replication fork while the other traps the anti-clockwise replication fork. Interestingly, both these proteins bind to DNA in a polar manner that blocks the replication fork in one direction, but allow it to continue in when approached from the other direction.
RTP
|
Structure of RTP
A single RTP monomer consists of four , three and an unstructured domain. The binds to DNA by into the major groove of Ter sites, while the interacts with the minor groove. The N-terminal arm also binds to the Ter site[1].
RTP conforms to the classic winged-helix motif, a compact α/β structure with αβααββ topology where "" project from the loop between and [2]. However, each monomer lacks a well-formed β1-strand (named β1-loop) and has an additional following the β3-strand[3].
()
RTP Polarity
The functional RTP-DNA complex requires two dimers of RTP. Dimerisation occurs through the extended of each monomer forming an anti-parallel coiled coil. The RTP dimer binds to the B site first which then facilitates cooperative binding of another RTP dimer to the A site to form a complete RTP-Ter complex[3].
It is thought that upon binding to the Ter sequence, the complex is able to impede the progress of replicative machinery headed by the replicative helicase. This is supported by in vitro studies involving E. coli where RTP was shown to interact with the DnaB helicase, thus a similar interaction may also occur in B. subtilis [4].
All of the Ter sites in the B. subtilis chromosome are only partially symmetric about the mid-point. However, initial studies of the RTP-DNA structure were performed using a symmetrically designed sequence (sRB) based on the TerI (the first Ter site that the clockwise replication fork encounters) B site [5]. NMR and crystallography revealed that RTP bound symmetrically to this symmetric sRB sequence. However, further studies using the non-symmetric native TerI B site (nRB) showed an asymmetric structure. Between the sRB and nRB sites there were only 6 base pair differences: three upstream and three downstream, but these changes were significant enough to influence how RTP binds to the DNA. Despite the change in sequence, the RTP-sRB interaction showed similar binding affinity to the RTP-nRB interaction, but had reduced fork arrest capability[5].
This differential capability fork arrest is due to the location of the 6 base pair changes. The three downstream changes have no bearing on the conformation of RTP as no base-specific interactions are made in this region. However, the three upstream changes are located within the major groove of the dsDNA of the TerI site, where the binds. Thus, it is these 3 bases that underlies the RTP binding specificity[3]. Changing these 3 crucial bases affected the binding of the RTP monomer to the upstream region and initiated the observed structural difference.
As RTP binds asymmetrically across the nRB, due to its differing binding affinities for the B and A sites, a polar complex is formed to block the replication fork. When the fork approaches the B site where there is tight RTP-DNA binding, the fork is unable to progress. But approaching from the A site, where there is a lower binding affinity, the fork is able to pass the complex.
RTP as a checkpoint regulator
An interesting feature is that RTP can cause replication fork pausing in B. subtilis at regions other than Ter sites, acting as a replication checkpoint regulator. There appear to be stringent terminator (STer) regions where RTP can bind and arrest replication when cells are under stress i.e. the stringent response. These regions are located around 200 kb either side of the origin of replication[6]. Studies have shown that there is 76% homology between this site and the regular Ter sites[7]. Additionally, mutational studies have reported that interaction between two dimers may not be required for arrest at STer sites. This is not the case with regular Ter sites.
Tus
|
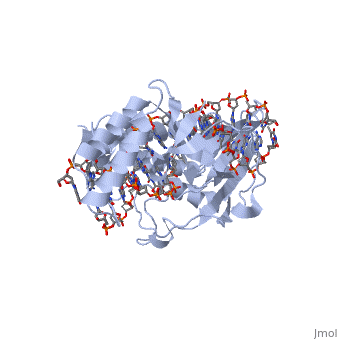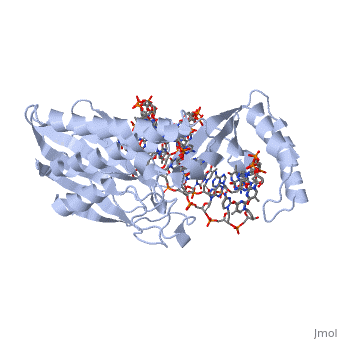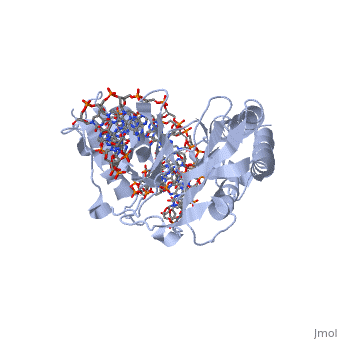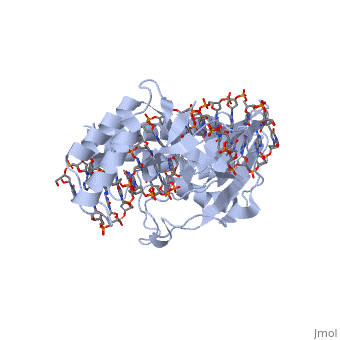
Structure of Tus
The Tus protein works as a monomer, each monomer consists of two domains, the amino and carboxy domains. The overall structure of Tus is made up of two separated by central twisted anti-parallel . These structures together form a that accommodates a B-form Ter . This binding involves two the bases and sugar-phosphate back bones from the major groove of the Ter DNA. Upon Tus binding, the major groove becomes deeper and the minor groove is significantly deeper, causing the DNA to be slightly bent.
()
Tus Polar Fork Arrest Mechanism
Like the RTP Ter sites, the Tus Ter sites do not display any symmetry or inverted repeats, thus a polar complex is formed between the Tus protein and Ter site. Despite both being replication termination proteins, RTP and Tus have very different structures and mechanisms of polar fork arrest.
The polar binding of Tus to its Ter site results in the protein having 2 different faces: a non-permissive face where the replication fork is blocked to terminate replication, and a permissive face where the replication machinery, headed by the DnaB helicase is allowed to pass and continue replication.
DnaB helicase normally functions by clamping onto only one of the single stranded DNA stands of the replication fork, and sliding in a 5’→3’ direction to progressively unwind the double stranded DNA ahead. It is this sliding movement of the DnaB helicase along the DNA that knocks Tus off its Ter binding site when approached from the permissive face of Tus that allows replication to continue. Tus dissociation occurs due to the progressive loss of Tus-Ter interactions.
On the other hand, when the DnaB helicase approaches from the non-permissive face, the DnaB helicase is unable to knock Tus off its Ter site due to the formation of a locked complex between Tus and it’s duplex Ter sequence.
Of particular importance to the formation of this complex is the bottom strand cytosine at position 6 of the Ter site (C6), which has been strictly conserved in all ten E. coli Ter sites. Formation begins with the DnaB helicase separating the Ter site DNA from the non-permissive face to release C6 from its base pair partner. This then leaves C6 to be freely base flipped out of the double helix, through rotation of the phosphodiester backbone, and into a pocket near the α4 helix of the Tus protein. Hydrogen bonding with Gly149, His144 and Leu150 positions C6 within the pocket. C6 is not the only base that undergoes change, the thymine at position 6 (T6) of the bottom strand is also moved out of the helix to allow C6 to reach to the pocket. T6 does not seem to make any base specific contacts with the Tus protein, as supported by the lack of conservation at this position of the Ter site.
The unlocking of the complex after it has performed it function of terminating replication from the non-permissive face, is ultimately required for the replication fork in the other direction to proceed. There is no specific unlocking mechanism, this is simply achieved as previously described, with the sliding movement of the DnaB helicase knocking Tus off its Ter sequence.
References
- ↑ Pai, S. K., Bussiere, D. E., Wang, F., Hutchinson, C. A., White, S. W. & Bastia, D. (1996) The structure and function of the replication terminator protein of Bacillus subtilis: identification of the ‘winged helix’ DNA-binding domain. EMBO J. 15(12), 3164-3173.
- ↑ Gajiwala, K. S. & Burley, S. K. (2000) Winged Helix proteins. Curr. Opin. Struct. Biol. 10, 110-116.
- ↑ 3.0 3.1 3.2 Vivian, J. P., Porter, C. J., Wilce, J. A. & Wilce, M. C. J. (2007) An Asymmetric Structure of the Bacillus subtilis Replication Terminator Protein in Complex with DNA. J. Mol. Biol. 370, 481-491.
- ↑ Gautam, A., Mulugu, S., Alexander, K. & Bastia, D. (2001) A Single Domain of the Replication Termination Protein of Bacillus subtilis Is Involved in Arresting Both DnaB Helicase and RNA Polymerase. J. Biol. Chem. 276(26), 23471-23479.
- ↑ 5.0 5.1 Wilce, J. A., Vivian, J. P., Hastings, A. F., Otting, G., Folmer, R. H., Duggin, I. G. et al. (2001) Structure of the RTP-DNA complex and the mechanism of polar replication fork arrest. Nature Struct. Biol. 8, 206-210.
- ↑ Levine, A., Vannier, F., Dehbi, M., Henckes, G. & Seror, S. J. (1991) The stringent response blocks DNA replication outside the ori region in Bacillus subtilis and at the origin in Escherichia coli. J. Mol. Biol. 219, 605-613.
- ↑ Autret, S., Levine, A., Vannier, F., Fujita, Y. & Seror, S. J. (1999) The replication checkpoint control in Bacillus subtilis: identification of a novel RTP-binding sequence essential for the replication fork arrest after induction of the stringent response. Mol. Microbiol., 31, 1665-1679.
