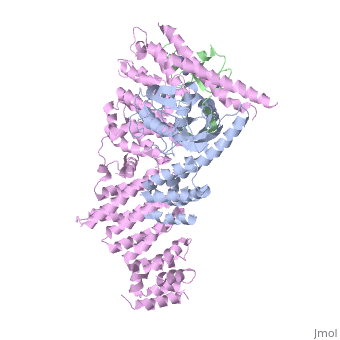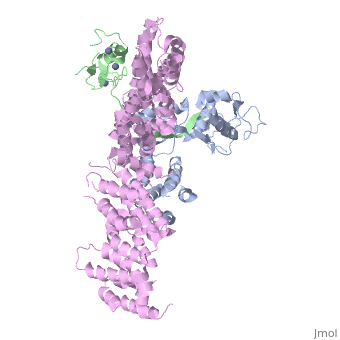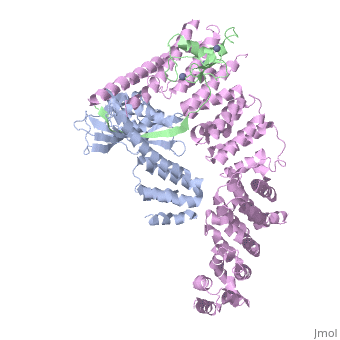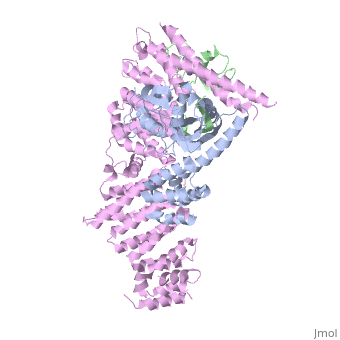This old version of Proteopedia is provided for student assignments while the new version is undergoing repairs. Content and edits done in this old version of Proteopedia after March 1, 2026 will eventually be lost when it is retired in about June of 2026.
Apply for new accounts at the new Proteopedia. Your logins will work in both the old and new versions.
4f52
From Proteopedia
Structure of a Glomulin-RBX1-CUL1 complex
Structural highlights
FunctionCUL1_HUMAN Core component of multiple cullin-RING-based SCF (SKP1-CUL1-F-box protein) E3 ubiquitin-protein ligase complexes, which mediate the ubiquitination of proteins involved in cell cycle progression, signal transduction and transcription. In the SCF complex, serves as a rigid scaffold that organizes the SKP1-F-box protein and RBX1 subunits. May contribute to catalysis through positioning of the substrate and the ubiquitin-conjugating enzyme. The E3 ubiquitin-protein ligase activity of the complex is dependent on the neddylation of the cullin subunit and is inhibited by the association of the deneddylated cullin subunit with TIP120A/CAND1. The functional specificity of the SCF complex depends on the F-box protein as substrate recognition component. SCF(BTRC) and SCF(FBXW11) direct ubiquitination of CTNNB1 and participate in Wnt signaling. SCF(FBXW11) directs ubiquitination of phosphorylated NFKBIA. SCF(BTRC) directs ubiquitination of NFKBIB, NFKBIE, ATF4, SMAD3, SMAD4, CDC25A, FBXO5 and probably NFKB2. SCF(SKP2) directs ubiquitination of phosphorylated CDKN1B/p27kip and is involved in regulation of G1/S transition. SCF(SKP2) directs ubiquitination of ORC1, CDT1, RBL2, ELF4, CDKN1A, RAG2, FOXO1A, and probably MYC and TAL1. SCF(FBXW7) directs ubiquitination of cyclin E, NOTCH1 released notch intracellular domain (NICD), and probably PSEN1. SCF(FBXW2) directs ubiquitination of GCM1. SCF(FBXO32) directs ubiquitination of MYOD1. SCF(FBXO7) directs ubiquitination of BIRC2 and DLGAP5. SCF(FBXO33) directs ubiquitination of YBX1. SCF(FBXO11) does not seem to direct ubiquitination of TP53. SCF(BTRC) mediates the ubiquitination of NFKBIA at 'Lys-21' and 'Lys-22'; the degradation frees the associated NFKB1-RELA dimer to translocate into the nucleus and to activate transcription. SCF(Cyclin F) directs ubiquitination of CP110 (By similarity).[1] [2] [3] [4] [5] Publication Abstract from PubMedThe approximately 300 human cullin-RING ligases (CRLs) are multisubunit E3s in which a RING protein, either RBX1 or RBX2, recruits an E2 to catalyze ubiquitination. RBX1-containing CRLs also can bind Glomulin (GLMN), which binds RBX1's RING domain, regulates the RBX1-CUL1-containing SCF(FBW7) complex, and is disrupted in the disease Glomuvenous Malformation. Here we report the crystal structure of a complex between GLMN, RBX1, and a fragment of CUL1. Structural and biochemical analyses reveal that GLMN adopts a HEAT-like repeat fold that tightly binds the E2-interacting surface of RBX1, inhibiting CRL-mediated chain formation by the E2 CDC34. The structure explains the basis for GLMN's selectivity toward RBX1 over RBX2, and how disease-associated mutations disrupt GLMN-RBX1 interactions. Our study reveals a mechanism for RING E3 ligase regulation, whereby an inhibitor blocks E2 access, and raises the possibility that other E3s are likewise controlled by cellular proteins that mask E2-binding surfaces to mediate inhibition. Structure of a Glomulin-RBX1-CUL1 Complex: Inhibition of a RING E3 Ligase through Masking of Its E2-Binding Surface.,Duda DM, Olszewski JL, Tron AE, Hammel M, Lambert LJ, Waddell MB, Mittag T, Decaprio JA, Schulman BA Mol Cell. 2012 Aug 10;47(3):371-82. Epub 2012 Jun 28. PMID:22748924[6] From MEDLINE®/PubMed®, a database of the U.S. National Library of Medicine. See Also
References
| ||||||||||||||||||